By Gertrude-Emilia Costin1 and Gopinathan K. Menon2
1Institute for In Vitro Sciences, Inc. (IIVS), Gaithersburg, MD, United States
2California Academy of Sciences, San Francisco, CA, United States
January 15, 2018
Cosmetic chemists best appreciate the implications of skin color for which the melanocytes are singularly responsible by producing melanin, the pigment which imparts the skin with its various shades of color. The marketing campaigns focused on the efficacy of personal care products to modulate skin color warrant an in-depth understanding of the melanocyte, its biology, and how influencing its functions changes skin appearance and health.
The Melanocyte: Definition and Origin
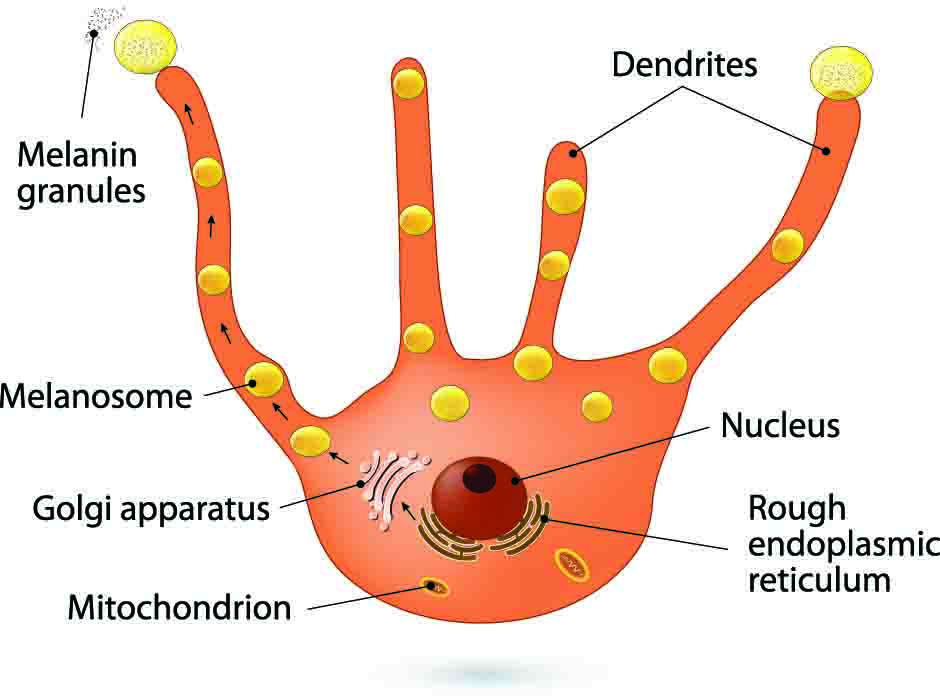
Melanocytes are defined as dendritic cells located in the basal layer of the epidermis (Figure 1). They produce the pigment melanin from tyrosine that is processed through a complex biosynthetic pathway governed by the rate limiting enzyme Tyrosinase (Figure 2) located within organelles termed melanosomes. Upon maturation, the melanin is transferred to keratinocytes for imparting pigmentation to skin.
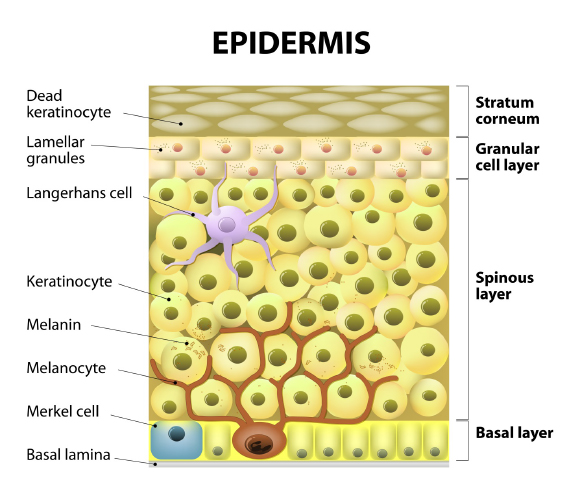
Figure 1: From the depth to the surface: melanocytes are located deep on the basal layer of skin and transfer the melanin they produce to the keratinocytes. Originally published in G.E. Costin, Decoding and modulating the color of human skin, TheCosmeticChemist.com, November 15, 2016.
Melanocytes originate from embryonic neuro-ectoderm, and are dispersed predominantly in the skin, but they can be found also in several other anatomical locations such as the cochlea of the ear, eye, adipose tissue, etc.1 There are profound reasons to be interested in melanocytes (in addition to cosmetic reasons) such as cultural and medical, as their products are the basis of racial and ethnic divisions and xenophobia that continues to shape human destiny. It must surprise the layman that whatever skin color they may have, everyone has the same number of melanocytes in their skin.2,3 Melanin offers protection from sun damage, and altered melanogenesis is the basis of seasonal adaptation. For the personal care industry, there is a huge opportunity to influence skin lightening or tanning, as the case may be, by modulating melanocyte function. For the healthcare field, melanoma continues to be a huge challenge, and disfiguring aspects of vitiligo are also a grave concern. Albinism among African tribals also have deadly social implications. Not surprisingly, the melanocyte (a cell that hides deep in the skin) possesses a very unique activity that ironically has more impactful implications globally than any other cell in the human body. The melanocytes are either "praised" for the beautiful skin color tones or "blamed" for racial discrimination.
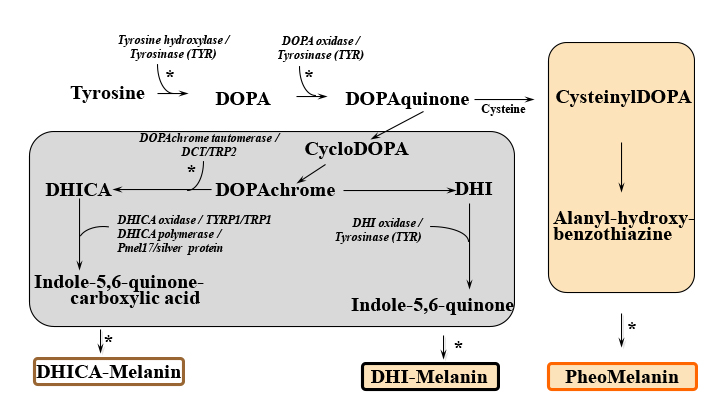
Figure 2: Melanin biosynthesis pathway. Abbreviations: DCT, Dopachrome tautomerase; DHI, 5,6-Dihydroxyindole; DHICA, DHI-2-carboxylic acid; DOPA, L-3,4-dihydroxyphenylalanine; TRP2, Tyrosinase related protein 2 (also known as DCT); TYR, Tyrosinase; TYRP1, Tyrosinase related protein 1 (also known as TRP1). In part, modified from: G.E. Costin, Decoding and modulating the color of human skin, TheCosmeticChemist.com, November 15, 2016.
Let us examine the intriguing story of the tiny, shy melanocyte at the crossroads of cultural politics of race, gender, and national attitudes of beauty and skin color. Today, the regimens of beauty include actives that do not drastically reduce the melanin content of the skin, but rather brighten the skin tone. Often a fine line separates the lightening of skin, which targets the reduction of pigmentation and focuses on producing an even skin tone, and the brightening of skin, which is directed to the restoration of vibrancy to the skin, with increased radiance and glow. Whichever may be the major consumer demand, the cosmetic industry finds itself in a race to innovate continuously while avoiding dramatic changes to skin’s homeostasis and addressing the needs of the consumer for green, organic products not tested on animals.
In Questions of Race, Ethnicity, and Xenophobia
Color tone in certain cultures relates to hierarchies of race, gender, caste, and ethnicity, which may not be immediately obvious to an outside observer. But scientists recognize that this is a matter of numbers (of melanosomes), concentrations (of melanin), or mutations that impact the biosynthesis of the pigment. This may sound rather cynical, but the reality of skin tone resides in microliters, picograms, and length of melanocyte dendrites, to say the least. It might surprise some people, but despite differences in ethnic groups or skin phototype, the number of melanocytes is the same, just the amount and type of melanin produced differs and leads to the wide array of skin tones. The amount of melanin produced is dependent on genetic factors and on the exposure to ultraviolet (UV) light.
The constitutive color of human skin varies widely and is frequently classified into one of six distinct skin phototypes.4 It is highly dependent on the type of melanin produced (pheomelanin or eumelanin) (Figure 2), maturity of the melanosomes (stages I to IV as shown in Figure 3a), and cellular events involved in the processing of the pigment.5 In lightly pigmented skin, primarily stage I and stage II melanosomes are reportedly found, whereas in darkly pigmented skin, stage IV melanosomes are predominant.6 The rate of melanosome degradation once they are transferred to the surrounding keratinocytes is an additional factor responsible for skin color differences and this is related in part to the size of the individual melanosomes. The smaller melanosomes of lightly pigmented skin have been found clustered in groups within secondary lysosomes (autophagosomes) in the keratinocytes and are degraded by the cells of the mid-stratum spinosum. In darkly pigmented skin, the melanosomes are larger and singly dispersed within lysosomes of the keratinocytes; their degradation is slower, and hence melanin granules can still be found in the stratum corneum. Furthermore, experiments using in vitro melanocyte cultures showed that the total melanin content was higher in a skin type VI melanocyte culture than in skin type I and II melanocyte cultures, thus confirming the clinical presentation of the main phototypes.7
Description of Melanocyte Morphology and Physiology
The specific function of the melanocyte is melanin synthesis within the organelles equipped to accomplish this task: the melanosomes. As the melanosomes mature within the cells, they gain the necessary enzymatic battery needed for the biosynthesis of melanin and start producing increasing amounts of the pigment. Progressing through stages I, II, and III, melanosomes become identifiable by transmission electron microscopy (Figure 3b). In the last stage of their development (stage IV), the melanosomes are fully melanized and are transported to surrounding keratinocytes by elements of the cytoskeletal system. One physiological characteristic of melanocytes is represented by their dendrites, an inheritance from their ancestors, the neural crests. Through their dendrites, the melanocytes establish close contact with the neighboring keratinocytes. Together they form the "epidermal melanin unit" usually composed of one melanocyte in contact with 35-40 neighboring keratinocytes, which regulate each other and contribute to the maintenance of skin health and homeostasis through secreted factors, receptors, and cell-cell contacts.8-10
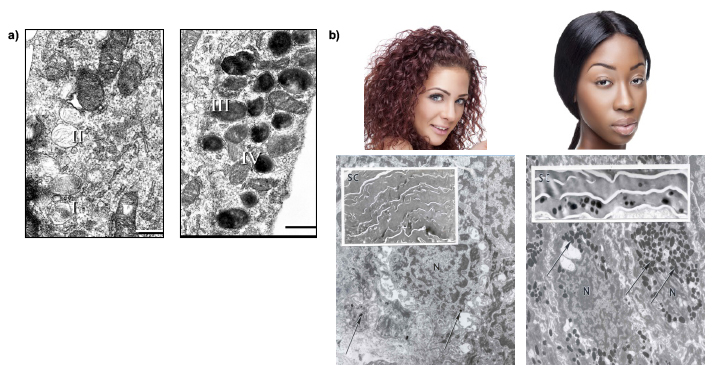
Figure 3: Race, ethnicity, and diversity. a) Stages of melanosome development. Images reproduced with permission (with slight modifications) from Costin et al. Tyrosinase processing and intracellular trafficking is disrupted in mouse primary melanocytes carrying the underwhite (uw) mutation. A model for oculocutaneous albinism (OCA) type 4, J. Cell. Sci., 116, 3203-3212 (2003); Copyright (2003) The Company of Biologists Ltd. b) Caucasian and African-American phototypes exemplified by appearance of skin and electron microscopy images. The presence of melanosome "microparasol" for nuclei in the basal layer is indicated by arrows; the insets indicate the presence of melanosomes in the stratum corneum.
The strictly regulated melanogenic pathway (Figure 2) leads to the formation of either pheomelanin or eumelanin depending on the availability of substrates and the specific function of the melanogenic enzymes involved in the process.11 Tyrosinase is the rate limiting enzyme of the pathway and catalyzes the first two steps: 1) the hydroxylation of tyrosine to L-3,4-dihydroxyphenylalanine (DOPA), which is 2) rapidly oxidized to DOPAquinone. When cysteine is available, DOPAquinone interacts with it producing 3- or 5-cysteinylDOPAs, which then oxidize, polymerize, and generate pheomelanin. In the absence of thiols, brown/black eumelanin is produced. For many years, Tyrosinase has been the most investigated factor of the pathway and most skin care actives in the field of pigment modulation were designed to affect its enzymatic activity. As research unraveled, newer aspects of pigmentation came to light and scientists (while not totally abandoning Tyrosinase as a target to modulate skin tone) have been more recently concentrating on other key points where modulators can affect the pathway, such as Tyrosinase processing, maturation, acquisition of enzymatic activity and degradation, and melanin transfer to keratinocytes.5,12 Other factors of the melanogenic pathway have been studied in various pigmentary conditions and may also be (or become) targets of modulators of interest to the cosmetic industry (Figure 2) though knowledge about subsequent effects is nowhere near that about Tyrosinase. Innovation resides in the minute changes that non-toxic compounds are capable to induce and subsequently affect events within the melanogenic pathway while not having devastating effects on the overall skin functionality.
Melanocytes are easily observed in culture using a simple inverted scope thanks to the presence of melanin pigments.13 They can also be detected using the classic Hematoxylin and Eosin staining that helps evaluate the physiological status of the cells. More specific or specialized staining procedures can be employed, such as the one using L-DOPA, a substrate of Tyrosinase that allows the detection of the enzyme either within the cells13 or in gels after western blotting.14 Positive expression of melanogenic proteins (Tyrosinase, Tyrosinase related protein-1, Dopachrome tautomerase, etc.) is also frequently used to demonstrate any effects compounds may have on these enzymes, and to provide support of their potential impact on the overall production of melanin.15 Fontana-Masson is a specialized stain for melanin, frequently used by researchers to investigate the distribution of melanin granules either in melanocyte cultures, skin explants, or reconstructed pigmented tissue models.16 All these assays are helpful to the cosmetic chemist in gaining valuable information about the possible mechanism of action of the molecules of interest. Shading light on the potential mechanism (or factors involved in it) provides a huge advantage in the claims made as part of the marketing presentation of finished products.
Types of Melanin and its Protective Roles
As mentioned above, there are two types of melanin in the human skin, the black/brown eumelanins and red/yellow pheomelanins, which exhibit distinct physical and biological properties.17,18 Eumelanin has a higher resistance to degradation and greater ability to neutralize reactive oxygen species (ROS) compared to pheomelanin, and therefore is attributed to photoprotection properties. As already discussed, despite the wide diversity of tones observed in the human race, the density of melanocytes in all types of skin is virtually identical.2,3 Therefore, we must admit that the differences in skin color depend on the amount of melanin produced, the efficiency of its transfer from melanocytes to keratinocytes, and the ratio of pheomelanin to eumelanin synthesized.19-21
Jablonski and Chaplin argue that during human evolution and dispersion across the globe, dark, photoprotective eumelanin pigmentation was selected as an adaptation to harsh UV radiation around the equator, considered to be of survival value to prevent loss of folic acid.22 In the low-UV regions, eumelanin and lighter skin evolved to allow for the synthesis of vitamin D in skin. While the limits of UV protection by melanin have been debated, it is generally accepted that the "microparasols" of melanin above the nuclei of basal epidermal layer (Figure 3b) provide a degree of protection to the DNA from UV radiation. Seasonal tanning of skin is also an indication of the protective value of melanin, as it is part of the skin’s interrelated defensive barriers.
Signaling Molecules, Hormones, and Cytokines that Modulate Melanocyte Function
The complexity of the melanogenic pathway relies on multiple factors acting in a highly regulated way to keep the process working within physiological parameters. Melanocytes are sensitive and highly responsive to their environment and modulate the levels of melanin production or proliferation according to extrinsic signals (such as UV) or intrinsic factors self-produced or derived from other cell types in the skin.10,23
As an extrinsic factor that skin comes in contact with constantly, UV has been investigated extensively and it is now known that it can act directly on melanocytes or can influence their activity by signaling through factors produced by keratinocytes. Following UV exposure, melanocytes over-express key enzymes involved in the melanogenic pathway, Tyrosinase,Tyrosinase related protein 1, and Dopachrome tautomerase.21,24 Furthermore, the oncogene p53 is over-expressed in keratinocytes, which directly stimulates the production of multiple factors that can then act in a paracrine manner to stimulate melanocyte function through receptor-mediated signaling pathways. Finally, UV facilitates the distribution of melanin to the supranuclear areas of the keratinocytes to protect them against DNA damage, and to the upper epidermis to protect the lower epidermis, which hosts melanocyte and keratinocyte stem cells.25
Among the intrinsic factors modulating the melanocyte functions, it is worth mentioning the following especially since some of them are key components of culture media for melanocyte cultures (discussed later in this article).
Factors synthesized in/secreted by melanocytes:
1. Enzymes involved in the melanogenic pathway and located in melanocytes (Figure 2).
2. Proteins with supportive roles in the melanin biosynthesis pathway: ocular albinism (OA)1 protein, P protein, membrane-associated transporter protein (MATP), copper-transporting P-type ATPase (ATP7A or Menkes protein) and biogenesis of lysosome-related organelles complex 1 (BLOC-1).26
3. Structural components within melanosomes: Pmel17/gp100/silver protein, whose sorting involves adaptor protein 1A (AP1A), AP1B, AP2, and spectrin, and chaperone-like component melanoma antigen recognized by T-cells 1 (MART-1), which plays structural roles in melanosome maturation.27-29
4. Factors implicated in melanosomes movement and transfer to keratinocytes: microtubules, dynein, kinesin, actin filaments, Rab27a, melanophilin, myosin Va, and Slp2-a.
5.
Transcription factors: among those, microphthalmia-associated transcription factor (MITF) plays a key role in the regulation of melanogenic proteins expression; other transcription factors of importance are paired box (PAX) 3, SRY-Box (SOX)9/10, and T-cell factor/lymphoid enhancer factor (TCF/LEF-1).
Factors secreted by other cell types of skin that influence the melanocytes:
1. Dermal fibroblasts: stem cell factor (SCF), neuregulin 1 (NRG1),30 or Dickkopf (DKK)1,31,32 which influence the growth and pigmentation of melanocytes, their shape, dendricity, mobility and adhesive properties.33
2. Endothelial cells: endothelin (ET-1), prostaglandins (PGs) and nitric oxide (NO).
3. Hormonal factors: that stimulate pigmentation include estrogen (which is related to pregnancy-induced pigmentation), alpha-melanocyte stimulating factor (MSH), adrenocorticotropic hormone (ACTH) and endorphin, while androgens have inhibitory effects on melanocytes.34
With so many possibilities to modulate the melanogenic pathway offered by numerous points of impact (Figure 2) and diverse cellular factors at their disposal, scientists face the dilemma of which one is safe to modulate while keeping in mind that minute changes can induce extreme effects in the appearance and physiology of skin. Fortunately, we have a battery of testing protocols of different levels of complexity ranging from cell cultures to skin explant-based assays. Since melanin has a primary protective role, any imbalance in its production (either reduction or stimulation) can have undesired and possible dangerous effects on skin and on the life of the affected individual. Therefore, before products are released on the market, industry members need to conduct extensive search of the literature and perform laboratory experiments to assess any adverse effects on skin and concurrently support the intended effect onto the melanogenic pathway.
The Melanocyte’s Response to Sun Exposure and its Implications in Photoaging
Melanocytes in skin respond to UV radiation resulting in: a) immediate pigment darkening within the first few minutes (and fading in a few hours) and b) delayed tanning that lasts relatively long.35 Immediate pigment darkening is thought to involve chemical modification (oxidation) of existing melanin and its precursors, and is seen in moderately pigmented to darkly pigmented skin. Its biological significance is still obscure.36
Delayed tanning is a longer term adaptation to sun exposure and has attracted more attention from researchers leading to more extensive studies. A detailed description of the process is not provided here (the reader is directed to the many references given at the end of the article), but some interesting findings on how the melanocyte senses UV and possibly other components of the solar spectrum to initiate the tanning response is of interest to cosmetic chemists. Several photoreceptor proteins—opsins—as well as their messenger RNAs have been identified in melanocytes (as well as in keratinocytes), which means that the skin can "see" UV radiation, blue light, etc. and respond appropriately.37 Besides, as mentioned in the foregoing sections, opiate receptors are also reported in the epidermis (which include keratinocytes and melanocytes). UV exposure leads to the release of endorphins from the epidermis (derived from metabolism of proopiomelanocortins), which would also kick start the tanning response. A recent study provided crucial evidence that exposure to chronic UV produced endogenous opioid dependence.38 In light of these recent discoveries, it is more important than ever for the scientific and medical community to educate the public that sunbathing and tanning beyond physiologically needed limits could lead to photoageing of skin and beyond, possibly leading to more dangerous outcomes such as melanoma.
Culturing Melanocytes: Knowing How To, and How Not To
When isolated from their natural environment and forced to in vitro cultures, melanocytes prove once more their delicate condition: they are rather demanding on conditions (pH, growth factors, splitting ratio and frequency, etc.) and like the company, especially that of keratinocytes. Melanocytes in the normal human epidermis are generally dendritic, do not proliferate, and do not express melanoma-associated antigens. Their physiology changes in culture, where they are bi- to tri-polar, proliferate with 2- to 4-day doubling times, and express melanoma-associated antigens. Keratinocytes and/or keratinocyte-derived factors are known to control growth, morphology, and antigen expression of normal melanocytes through cell-cell contact or by addition to the culture media, respectively. Early studies showed that melanocytes separated from keratinocytes by semiporous membranes remained di- and tri-polar and could not proliferate in medium optimal for keratinocytes. When cell-cell contact was established between melanocytes and undifferentiated keratinocytes, melanocytes proliferated at a rate similar to keratinocytes and they developed multiple dendrites. In co-cultures, melanocytes proliferated when surrounded by differentiated keratinocytes in the intermediate and upper layers.39 These early studies proved critical in understanding the dynamics of melanocytes cultured in vitro and support the paradigm that the closer they are brought to their native environment (co-cultures with keratinocytes or reconstructed tissue models), the better chance the scientist has for accuracy of results and reliable interpretation, leading to sound conclusions.
Melanocytes are adherent cells, therefore using well established protocols is critical as otherwise they may not grow (especially the human lines which grow slower compared to mouse lines) or affect the production of melanin at a normal, natural rate. Furthermore, the melanocytes have low mitotic activity compared to other skin resident cells and are usually rapidly overwhelmed in culture by keratinocytes and fibroblasts.40 This challenge was overcome by the use of tumor promoting phorbol ester 12-O-tetradecanoyl phorbol-13-acetate (TPA), which enhances the proliferation and attachment of melanocytes and is toxic to keratinocytes.41 Later on, geneticin (G418) was used to eliminate the fibroblasts from culture and became a critical first step in raising pure melanocyte cultures from resected skin samples.42 Other additives used in cultures of melanocytes are cyclic AMP(cAMP)-inducers, such as cholera toxin (CT) and isobutyl-methylxanthine (IBMX), which have a mitogenic effect on melanocytes and promote their division. Keratinocyte-derived growth factors such as basic fibroblast growth factor (bFGF), nerve growth factor (NGF), and endothelin-1 (ET-1) play key roles in cultures of melanocytes, enhancing their proliferation, Tyrosinase activity, and dendricity.43
Experiments can be conducted with primary or immortalized cell lines of human or animal origin, or with melanoma lines, which tend to be easier to culture, at least from the standpoint of proliferation. Complex protocols have been developed to allow immortalization of melanocytes carrying various mutations in the melanogenic proteins, some of them making the immortalization a challenging if not impossible task.44 Even though in many laboratories culturing skin melanocytes has become a routine research activity implying familiarity with regular culturing practices, investigations have revealed that the quality and quantity of the pigment formed in the cells cultured in static conditions (no changes in the media or treatment with actives) may differ significantly from those of the original skin pigment cells.7 In this study, conducted by Smit and coworkers, they showed that there were notable passage-to-passage variations in the synthesis of melanin, especially for pheomelanin. Therefore, it becomes imperative to analyze the melanin in the cells before the start of experiments and to troubleshoot the protocols such that the native pheno- and photo-type of the melanocytes is maintained throughout experiments to warrant reliable results.
It seems that each disease or group of medical conditions has its own celebrity cell line. Say "cervical cancer" and HeLa cells come to mind immediately. This is probably the oldest and most commonly used human cell line that was reportedly used in an impressive number of 96,754 manuscripts published and indexed in PubMed as of December 2017. Not as famous as HeLa, but in the realm of skin, are the HaCaT cells, a spontaneously transformed immortal keratinocyte line from adult skin often used in skin focused experiments. Now say "pigmentation" and anyone in the field will start sharing about experiments performed with the B16 melanoma line, discovered and maintained in the Jackson Labs since 1954 (Figure 4) and famous in its own right judging by an impressive number of 15,243 manuscripts indexed in PubMed. Since it is relatively easy to culture and proliferate, the B16 line is used as a to-go approach to investigate effects of actives on melanin production. Pigment synthesis by mouse melanoma cells is very sensitive to the culture conditions, especially cell density, growth rate, tyrosine concentration, and pH. For long term maintenance, it is advisable to grow the cells under non-pigmenting conditions and allow them to multiply as usual, by ~20 fold in 3 days. For pigmentation, the media can be changed to contain higher tyrosine concentration, higher pH, alpha-MSH, etc. However, if grown for a long time in media facilitating pigmentation, the melanocytes initially become pigmented, but after a while the production slows down the growth. Eventually, any cell not capable to produce pigment will overtake the culture and thus become selected, leading to a culture unable to produce melanin. Great resources of established protocols reside with the Welcome Trust Fund Genomics Cell Bank, Lab of Dr. Sviderskaya and Dr. Bennett who also hold repositories of various valuable lines of normal melanocytes or melanoma lines of several species.45,46 Another repository of skin specimens and skin lines (including melanomas) reside at Yale University (Specimen Resource Core within SPORE).47 The efforts put into making these resources available to the scientific community and their maintenance should be commended as they reflect the work of a scientific lifetime.
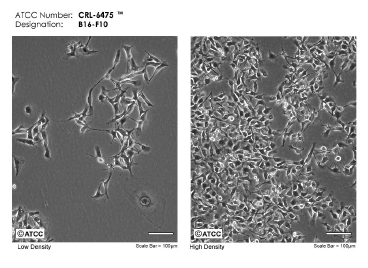
Figure 4: From skin to the cell culture flask. B16 mouse melanoma cells in culture at low and high density. Micrographs of B16-F10 (ATCC® CRL-6475) courtesy of ATCC.
Signs of incorrect culturing techniques are loss of pigmentation capacity and dendricity. Those are also the signs that the scientist will look for or desire when hoping to discover the next generation of efficacious actives impacting the skin tone as long as they are not related to toxicity, but rather to a specific impact on factors involved in the melanogenic pathway and beyond.
Just like any other cell in the human body, melanocytes are not immune to cancerous transformation. Among well-established intrinsic risk factors for developing melanoma are family history, multiple moles, fair skin, blue eyes, red hair, and freckles.48 Extrinsic factors including UV exposure and indoor tanning have also been associated with increased melanoma risk.49,50 Exposure to the Sun has been long determined be the responsible factor for the appearance of melanoma. Despite advances in its detection and treatment, melanoma is still the primary cause of mortality from skin disease in the Western world.51,52
And, you may ask why would the cosmetic chemist be interested in melanoma? In light of the increasing incidence for UV-induced skin cancer and the progressive depletion of the ozone layer, gaining a better understanding of the role of melanin in preventing UV-induced DNA damage and malignant transformation of skin cells is critical as the cosmetic chemist deals with a delicate target (the melanocyte) and a double edged sword molecule (melanin) when attempting to modulate skin tone. As we stressed throughout this article, careful, highly controlled changes in the melanogenic pathway are to be conducted, documented, and demonstrated for any actives with potential to affect the outcome of a melanocyte's work. If the imbalance is significant, the turnover to melanoma may be an obvious outcome, the worst case scenario that no one would like to be part of as a scientist. To further improve your understanding of the factors contributing to melanoma, we leave you with the reference by Chang and coworkers, suggestively entitled "More skin, more sun, more tan, more melanoma".53
We also take this opportunity to comment on the value of scientific societies such as the PanAmerican Society for Pigment Cell Research (PASPCR) and its counterparts from other geographic regions to share science trends and advancements in raising awareness on the medical and social implications of pigmentary disorders. Take for example the 2016 April issue of the PASPCR Newsletter where we can find information about the European Days of Albinism, Vitiligo International Symposium, and the impressive article by Don Sawatzky entitled "Under the same sun", in which he shares with us the efforts to protect people with albinism in Africa, to raise awareness about the atrocities against them, and to find ways of alleviating their conditions through scientific advances.54 By these venues, scientific community advocates for people impacted by pigmentary conditions, strives to find cures and treatments, and to voice social implications of these disorders with devastating effects on patients’ lives.
In this article, we traveled deep into the skin to discover the world of the melanocyte and bring its message to the surface. It seems that whether we are on the lookout for a gentle skin lightening active, in search for that miracle anti-ageing cream capable of stealing years away, or on the hunt for the natural/non-UV tanning procedure, the melanocytes hold the key. But they should not be taken for granted, as if after ending their treatment with compounds affecting their capacity to produce melanin, they would easily and immediately revert to their natural condition without any remnant effects. The plasticity of melanocytes is very tempting, but at the same time they are very sensitive cells, highly responsive to stimuli and easily susceptible to mutations or dysregulations that can dramatically change one's appearance, which could also have implications in their social and personal life. All scientists involved in attempts to modulate skin tone must exercise highly documented diligence and base the content of any product on high quality experiments and rationales.
1. M. Brenner and V.J. Hearing, What are the melanocytes really doing all day long...?: from the viewPoint of a keratinocyte: melanocytes – cells with a secret identity and incomparable abilities, Exp. Dermatol., 18, 799-819 (2009).
2. Y. Yamaguchi and V.J. Hearing, Melanocyte distribution and function in human skin: effects of UV radiation. In: From Melanocytes to Malignant Melanoma, Eds. V.J. Hearing and S.P.L. Leong, Humana Press: Totowa, 101-116 (2005).
3. Y.
Yamaguchi, K. Takahashi, B.Z. Zmudzka, A. Kornhauser, S.A. Miller, T. Tadokoro, W. Berens, J.Z. Beer, and V.J. Hearing, Human skin responses to UV radiation: pigment in the upper epidermis protects against DNA damage in the lower epidermis and facilitates apoptosis, FASEB J., 20, 1486-1488 (2006).
4. T.B. Fitzpatrick, The validity and practicability of sun-reactive skin types I through VI, Arch. Dermatol., 124, 869-871 (1988).
5. G.E. Costin, Decoding and modulating the color of human skin, TheCosmeticChemist.com, November 15, 2016.
6. J.L.
Bolognia and J.M. Pawelek, Biology of hypopigmentation, J. Am. Acad. Dermatol., 19, 217-235 (1988).
7. N.P. Smit, R.M. Kolb, E.G. Lentjes, K.C. Noz, H. van der Meulen, H.K. Koerten, B.J. Vermeer, and S. Pavel, Variations in melanin formation by cultured melanocytes from different skin types, Arch. Dermatol. Res., 290, 342-349 (1998).
8. Z. Abdel-Malek, V.B. Swope, and J.J. Nordlund, The nature and biological effects of factors responsible for proliferation and differentiation of melanocytes, Pigment Cell Res., 3 Suppl. 2, 43-47 (1992).
9. Z.A. Abdel-Malek, V.B. Swope, J. Pallas, K. Krug, and J.J. Nordlund, Mitogenic, melanogenic, and cAMP responses of cultured neonatal human melanocytes to commonly used mitogens, J. Cell. Physiol., 150, 416-425 (1992).
10. R. Halaban, The regulation of normal melanocyte proliferation, Pigment Cell Res., 13, 4-14 (2000).
11. S. Ito and K. Wakamatsu, Chemistry of melanins. In: The Pigmentary System. Physiology and Pathophysiology, Eds. J.J. Nordlund, R.E. Boissy, V.J. Hearing, R.A. King, W.S. Oetting, and J.P. Ortonne, Blackwell Publishing: Oxford, UK, 282-310 (2006).
12. G.E.
Costin and M. Randhawa, Skin tone modulation: in vitro pre-clinical and clinical efficacy testing strategies and innovative solutions in cosmetics, webinar by Institute for In Vitro Sciences and Johnson & Johnson, June, 29, 2017.
13. J. Tang, Q. Li, B. Cheng, and L.J. Jing, Primary culture of human face skin melanocytes for the study of hyperpigmentation, Cytotechnology, 66, 891-898 (2014).
14. S.M. Petrescu, A.J. Petrescu, H.N. Titu, R.A. Dwek, and F.M. Platt, Inhibition of N-glycan processing in B16 melanoma cells results in inactivation of tyrosinase but does not prevent its transport to the melanosome, J. Biol. Chem., 272, 15796-15803 (1997).
15. H. Watabe, J.C. Valencia, E. Le Pape, Y. Yamaguchi, M. Nakamura, F. Rouzaud, T. Hoashi, Y. Kawa, M. Mizoguchi, and V.J. Hearing, Involvement of dynein and spectrin with early melanosome transport and melanosomal protein trafficking, J. Invest. Dermatol., 128, 162-174 (2008).
16. N.
Mujahid, Y. Liang, R. Murakami, H.G. Choi, A.S. Dobry, J. Wang, Y. Suita, Q.Y. Weng, J. Allouche, L.V. Kemeny, A.L. Hermann, E.M. Roider, N.S. Gray, and D.E. Fisher, A UV-independent topical small-molecule approach for melanin production in human skin, Cell Rep., 19, 2177-2184 (2017).
17. G. Prota, Melanins and Melanogenesis, Academic Press: New York (1992).
18.
K. Ito, K. Wakamatsu, and H. Ozeki, Chemical analysis of melanins and its application to the study of the regulation of melanogenesis, Pigment Cell Res., 13, 103-109 (2000).
19. J.L.
Rees, The genetics of sun sensitivity in humans, Am. J. Hum. Gen., 75, 739-751 (2004).
20. K. Wakamatsu, R. Kavanagh, A.L. Kadekaro, S. Terzieva, R.A. Sturm, S. Leachman, Z. Abdel-Malek, and S. Ito, Diversity of pigmentation in cultured human melanocytes is due to differences in the type as well as quantity of melanin, Pigment Cell Res., 19, 154-162 (2006).
21. Y. Miyamura, S.G. Coelho, R. Wolber, S.A. Miller, K. Wakamatsu, B.Z. Zmudzka, S. Ito, C. Smuda, T. Passeron, W. Choi, J. Batzer, Y. Yamaguchi, J.Z. Beer, and V.J. Hearing, Regulation of human skin pigmentation and responses to ultraviolet radiation, Pigment Cell Res., 20, 2-13 (2007).
22. N.G. Jablonski and G. Chaplin, Colloquium paper: human skin pigmentation as an adaptation to UV radiation, Proc. Natl. Acad. Sci. U.S.A., 107, 8962-8968 (2010).
23. R. Busca and R. Ballotti, Cyclic AMP: A key messenger in the regulation of skin pigmentation, Pigment Cell Res., 13, 60-69 (2000).
24.
T. Tadokoro, Y. Yamaguchi, J. Batzer, S.G. Coelho, B.Z. Zmudzka, S.A. Miller, R. Wolber, J.Z. Beer, and V.J. Hearing, Mechanisms of skin tanning in different racial/ethnic groups in response to ultraviolet radiation, J. Invest. Dermatol., 124, 1326-1332 (2005).
25. T.S. Kuroda and M. Fukuda, Rab27A-binding protein Slp2-a is required for peripheral melanosome distribution and elongated cell shape in melanocytes, Nat. Cell Biol., 6, 1195-1203 (2004).
26. Y. Yamaguchi and V.J. Hearing, Physiological factors that regulate skin pigmentation, Biofactors, 35, 193-199 (2009).
27. T. Kushimoto, J.C. Valencia, G.E. Costin, K. Toyofuku, H. Watabe, K. Yasumoto, F. Rouzaud,W.D. Vieira, and V.J. Hearing, The Seiji memorial lecture: the melanosome: an ideal model to study cellular differentiation, Pigment Cell Res., 16, 237-244 (2003).
28. K.
Yasumoto, H. Watabe, J.C. Valencia, T. Kushimoto, T. Kobayashi, E, Appella, and V.J. Hearing, Epitope mapping of the melanosomal matrix protein gp100 (PMEL17): rapid processing in the endoplasmic reticulum and glycosylation in the early Golgi compartment, J. Biol. Chem., 279, 28330-28338 (2004).
29. T.
Hoashi, H. Watabe, J. Muller, Y. Yamaguchi, W.D. Vieira, and V.J. Hearing, Mart-1 is required for the function of melanosomal matrix protein PMEL17/GP100 and the maturation of melanosomes, J. Biol. Chem., 280, 14006-14016 (2005).
30. W. Choi, L. Kolbe, and V.J. Hearing, Characterization of the bioactive motif of neuregulin-1, a fibroblast-derived paracrine factor that regulates the constitutive color and the function of melanocytes in human skin, Pigment Cell Melanoma Res., 25, 1-5 (2012).
31.
Y. Yamaguchi, S. Itami, H. Watabe, K. Yasumoto, Z.A. Abdel-Malek, T. Kubo, F. Rouzaud, A. Tanemura, K.Yoshikawa, V.J. Hearing, Mesenchymal-epithelial interactions in the skin: increased expression of dickkopf1 by palmoplantar fibroblasts inhibits melanocyte growth and differentiation, J. Cell Biol., 165, 275-285 (2004).
32. Y.
Yamaguchi, T. Passeron, T. Hoashi, H. Watabe, F. Rouzaud, K. Yasumoto, T. Hara, C. Tohyama, I. Katayama, T. Miki, and V.J. Hearing, Dickkopf 1 (DKK1) regulates skin pigmentation and thickness by affecting Wnt/β-catenin signaling in keratinocytes, FASEB J., 22, 1009-1020 (2008).
33. G.E.
Costin and V.J. Hearing, Human skin pigmentation: melanocytes modulate skin color in response to stress, FASEB J., 21, 976-994 (2007).
34. T.
Tadokoro, F. Rouzaud, S. Itami, V.J. Hearing, and K. Yoshikawa, The inhibitory effect of androgen and sex-hormone-binding globulin on the intracellular cAMP level and tyrosinase activity of normal human melanocytes, Pigment Cell Res., 16, 190-197 (2003).
35. J. Moan, K.P. Nielsen, and A. Juzeniene, Immediate pigment darkening: its evolutionary roles may include protection against folate photosensitization, FASEB J., 26, 3971-3975 (2012).
36.
C. Routaboul, A. Denis, and A. Vinche, Immediate pigment darkening: description, kinetic and biological function, Eur. J. Dermatol., 9, 95-99 (1999).
37. K. Haltaufderhyde, R.N. Ozdeslik, N.L. Wicks, J.A. Najera, and E. Oancea, Opsin expression in human epidermal skin, Photochem. Photobiol., 91, 117-123 (2015).
38.
G.L. Fell, K.C. Robinson, J. Mao, C.J. Woolf, and D.E. Fisher, Skin beta endorphin mediates addiction to UV light, Cell, 157, 1527-1534 (2014).
39. I.T. Valyi-Nagy, G. Hirka, P.J. Jensen, I.M. Shih, I. Juhasz, and M. Herlyn, Undifferentiated keratinocytes control growth, morphology, and antigen expression of normal melanocytes through cell-cell contact, Lab Invest., 69, 152-159 (1993).
40. M.R. Pittelkow and G.D. Shipley, Serum-free culture of normal human melanocytes: growth kinetics and growth factor requirements, J. Cell. Physiol., 140, 565-576 (1989).
41. M. Eisinger and O. Marko, Selective proliferation of normal human melanocytes in vitro in the presence of phorbol ester and cholera toxin, Proc. Natl. Acad. Sci. USA, 79, 2018-2022 (1982).
42. R. Halaban and F.D. Alfano, Selective elimination of fibroblasts from cultures of normal human melanocytes, In Vitro, 20, 447-450 (1984).
43. M.
Akio, I. Hajime, O. Hideo, and K. Norio, Tyrosinase induction in normal human cultured melanocytes by endothelin-1, J. Cardivasc. Pharmacol., 44, S439-S442 (2004).
44. G.E. Costin, W.D. Vieira, J.C. Valencia, F. Rouzaud, L. Lamoreux, and V.J. Hearing, Immortalization of mouse melanocytes carrying mutations in various pigmentation genes, Anal. Biochem., 335, 171-174 (2004).
45.
https://www.sgul.ac.uk/depts/anatomy/pages/WTFGCB.htm
46. https://www.sgul.ac.uk/depts/anatomy/pages/dcbm&m.htm
47. http://medicine.yale.edu/dermatology/spore/
48. J.M. Elwood and J. Jopson, Melanoma and sun exposure: an overview of published studies, Int. J. Cancer, 73, 198-203 (1997).
49. W. Ting, C.K. Schultz, N.N. Cac, M. Peterson, and H.W. Walling, Tanning bed exposure increases the risk of malignant melanoma, Int. J. Dermatol., 46, 1253-1257 (2007).
50. M. Zhang, A.A. Qureshi, A.C. Geller, L. Frazier, D.J. Hunter, and J. Han, Use of tanning beds and incidence of skin cancer, J. Clin. Oncol., 30, 1588-1593 (2012).
51. A. Jemal, S.S. Devesa, P. Hartge, and M.A. Tucker, Recent trends in cutaneous melanoma incidence among whites in the United States, J. Natl. Cancer Inst., 93, 683-687 (2001).
52.
Á. Flórez and M. Cruces, Melanoma epidemic: true or false? Int. J. Dermatol., 43, 405-407 (2004).
53. C. Chang, E.C. Murzaku, L. Penn, N.R. Abbasi, P.D. Davis, M. Berwick, and D. Polsky, More skin, more sun, more tan, more melanoma, Am. J. Public Health, 104, e92-e99 (2014).
54. PASPCR Newsletter, April 2016.